夏季熱應(yīng)激是導(dǎo)致泌乳奶牛生育能力低下的主要因素之一。熱應(yīng)激導(dǎo)致母體體溫過高�����,生殖道細(xì)胞出現(xiàn)各種功能障礙���,生育能力低下。研究表明���,GV期階段卵母細(xì)胞發(fā)育能力受損是夏季生育能力低下的原因之一�����。研究人員在夏季時(shí)從中等大小的竇卵泡(直徑2-8毫米)收集的卵母細(xì)胞����,在體外受精或化學(xué)激活孤雌生殖后的發(fā)育能力低于涼爽季節(jié)的卵母細(xì)胞��。雖然早期竇卵泡階段是卵母細(xì)胞獲得發(fā)育能力的關(guān)鍵階段�����,但目前還沒有關(guān)于熱應(yīng)激對(duì)早期竇卵泡生長影響的報(bào)道���。
來自竇卵泡(2-8 mm)的GV期熱應(yīng)激卵母細(xì)胞的低發(fā)育能力與膜脂肪酸譜的變化增加及參與卵子發(fā)生���、卵泡發(fā)生和胚胎發(fā)育的基因轉(zhuǎn)錄水平的改變有關(guān)。暴露于體外高溫或夏季收集的GV期卵母細(xì)胞也表現(xiàn)出細(xì)胞核和細(xì)胞質(zhì)的破壞�,包括皮質(zhì)顆粒易位至卵膜,以及成熟卵母細(xì)胞中線粒體分布和極化受損���。然而��,GV期卵母細(xì)胞在生長過程中發(fā)育能力受到破壞的機(jī)制仍不清楚��。
發(fā)育卵泡中類固醇生成的變化可能與夏季卵母細(xì)胞發(fā)育能力降低有關(guān)�。類固醇激素是卵泡生長過程中卵母細(xì)胞獲得發(fā)育能力所需的重要因素��。在體外�����,雌二醇(E2)對(duì)于在早期竇卵泡(直徑0.4-0.7毫米)衍生的牛卵母細(xì)胞復(fù)合物的體外培養(yǎng)過程中獲得成熟和發(fā)育能力至關(guān)重要���。體內(nèi)研究表明�����,在高溫季節(jié)時(shí)發(fā)情周期的第7天���,優(yōu)勢(shì)卵泡的卵泡液中E2濃度低于低溫季節(jié)��。各種大小的竇卵泡的類固醇生成能力也會(huì)受到熱應(yīng)激的破壞���。在急性熱應(yīng)激后3周,從中等大小的卵泡(直徑6-9毫米)獲得的培養(yǎng)顆粒和膜細(xì)胞中E2和雄烯二酮(A4)的產(chǎn)生水平降低��,表明早期竇卵泡對(duì)熱應(yīng)激敏感��。尚未直接研究在熱應(yīng)激影響下生長卵泡中的類固醇生成與卵母細(xì)胞發(fā)育能力之間的關(guān)系���。
高溫誘導(dǎo)的氧化應(yīng)激被認(rèn)為是熱應(yīng)激損害卵母細(xì)胞發(fā)育能力的另一種潛在機(jī)制�。體外成熟系統(tǒng)的研究表明����,熱暴露會(huì)增加牛卵母細(xì)胞中的細(xì)胞內(nèi)ROS水平,并降低囊胚率�����。谷胱甘肽是哺乳動(dòng)物細(xì)胞中含量最豐富的非蛋白質(zhì)硫醇�,還原型谷胱甘肽(GSH)可維持細(xì)胞氧化還原狀態(tài),并保護(hù)細(xì)胞免受ROS侵害���。顆粒細(xì)胞具有在通過跨帶突起(TZP)的胞間連接向卵母細(xì)胞提供GSH的關(guān)鍵作用����。卵母細(xì)胞中細(xì)胞內(nèi)GSH水平的降低伴隨著IVM期間熱暴露引起的細(xì)胞內(nèi)ROS水平的增加�。在IVM培養(yǎng)基中添加半胱氨酸可刺激GSH合成,降低卵母細(xì)胞中的ROS水平并減輕熱暴露對(duì)卵母細(xì)胞發(fā)育能力的負(fù)面影響��。
一項(xiàng)研究旨在通過給奶牛喂食具有抗氧化能力的不飽和脂肪酸來提高夏季奶牛的卵母細(xì)胞質(zhì)量����。然而,喂養(yǎng)過程并沒有提高收集到的卵母細(xì)胞的發(fā)育能力���。如果我們可以通過使用IVG培養(yǎng)系統(tǒng)開發(fā)一個(gè)模擬熱應(yīng)激下卵泡生長的實(shí)驗(yàn)?zāi)P?����,評(píng)估補(bǔ)充某些物質(zhì)(即抗氧化劑)對(duì)生長期受熱應(yīng)激的卵母細(xì)胞發(fā)育能力的影響可能更容易����。實(shí)驗(yàn)?zāi)P椭羞x擇的有效物質(zhì)可用于在夏季管理奶牛����,為IVM(體外成熟)收集更優(yōu)質(zhì)的卵母細(xì)胞��,并在隨后的秋季提高生育能力���。
在體內(nèi),季節(jié)性熱應(yīng)激可能不僅會(huì)通過高溫?fù)p害卵母細(xì)胞在生長期的發(fā)育能力�����,也可能通過熱應(yīng)激環(huán)境下的其他生理變化(即促性腺激素分泌受損����,由于采食量減少導(dǎo)致的代謝系統(tǒng)改變等)。而在本研究中���,我們旨在使用牛卵母細(xì)胞的IVG培養(yǎng)系統(tǒng)����,研究生理范圍內(nèi)的高溫對(duì)GV期卵母細(xì)胞在生長期的影響�����。IVG培養(yǎng)使來自早期竇卵泡(直徑0.5-1毫米)的沒有成熟能力的卵母細(xì)胞能夠生長并獲得成熟和發(fā)育能力以發(fā)育到囊胚階段��。首先,我們檢查了IVG培養(yǎng)過程中熱暴露對(duì)囊胚階段的生長�����、成熟和隨后的發(fā)育能力的影響��。其次��,為了研究熱暴露降低GV期卵母細(xì)胞發(fā)育能力的機(jī)制���,我們?cè)u(píng)估了顆粒細(xì)胞產(chǎn)生的E2和孕酮 (P4),卵母細(xì)胞的細(xì)胞內(nèi)ROS和GSH水平���,以及卵母細(xì)胞和卵丘細(xì)胞之間的TZP數(shù)量�。第三�,我們研究了在IVG培養(yǎng)基中添加半胱氨酸對(duì)GSH水平的拯救作用以及在IVG期間暴露于高溫的卵母細(xì)胞的發(fā)育能力。
實(shí)驗(yàn)1:
IVG培養(yǎng)過程中熱暴露對(duì)OCGC形態(tài)����、卵母細(xì)胞生長以及卵母細(xì)胞成熟和發(fā)育能力的影響
為了檢查熱暴露對(duì)卵母細(xì)胞生長、成熟和發(fā)育能力的影響�,我們?cè)趦煞N不同的溫度設(shè)置(對(duì)照組和熱休克組;圖1a)下對(duì)源自早期竇卵泡的牛OCGC進(jìn)行了IVG���。在對(duì)照組中����,OCGC在38.5℃下培養(yǎng)12天,模擬非熱應(yīng)激奶牛的體溫�����。在熱休克組中�,OCGCs采用38.5℃ 5小時(shí)、39.5℃ 5小時(shí)��、40.5℃ 5小時(shí)和39.5℃ 9小時(shí)12天的溫度循環(huán)培養(yǎng)����,模擬熱應(yīng)激下奶牛的體溫。在IVG培養(yǎng)后�����,對(duì)卵母細(xì)胞進(jìn)行IVM�����、IVF和胚胎體外培養(yǎng)(IVC)�����,評(píng)估卵母細(xì)胞的直徑、核成熟能力和發(fā)育能力(圖1b)���。根據(jù)受精卵母細(xì)胞的數(shù)量計(jì)算囊胚率���。在IVG培養(yǎng)期間��,我們觀察了兩組OCGC的形態(tài)學(xué)表現(xiàn)(圖2)�,但對(duì)照組和熱休克組之間OCGC的活力或腔形成率沒有顯著差異(圖3a、b)����。組別和培養(yǎng)期的卵母細(xì)胞的平均直徑有顯著差異(P < 0.001)(圖3c)。此外��,組別和培養(yǎng)期對(duì)卵母細(xì)胞平均直徑有顯著的交互作用(P < 0.001)�����。對(duì)照組卵母細(xì)胞的平均直徑從IVG前的101.4 μm增加到IVG和IVM后的113.5 μm (P < 0.05)�。熱休克組直徑的增加也很顯著(從101.4到109.4 μm,P < 0.05)���,而熱休克組IVG和IVM后卵母細(xì)胞的平均直徑明顯小于對(duì)照組(P < 0.05)�。對(duì)照組(分別為62.1%和55.3%)和熱休克組(分別為51.9%和45.8%)之間的核成熟和分裂率相似(圖3c,表1)����。然而,熱休克組沒有卵母細(xì)胞發(fā)育成囊胚(0.0%)�����,而對(duì)照組有27.7%的卵母細(xì)胞發(fā)育成囊胚(P < 0.05)(表1)�。

圖1.對(duì)照組和熱休克組體外生長(IVG)培養(yǎng)過程中的溫度條件(a)和實(shí)驗(yàn)設(shè)計(jì)示意圖(b)。(a) 熱應(yīng)激下泌乳奶牛直腸溫度的每日變化(實(shí)線)�,以及熱休克組(實(shí)心正方形)和對(duì)照(空心圓)的培養(yǎng)溫度,由放置在孵化器中的數(shù)據(jù)記錄器監(jiān)測(連續(xù)12天每小時(shí)測量的平均溫度)���。熱休克組中的卵母細(xì)胞-卵丘-顆粒復(fù)合物(OCGC)在與熱應(yīng)激奶牛所經(jīng)歷的溫度相似的溫度范圍內(nèi)培養(yǎng)���。對(duì)照組的OCGCs在38.5℃的恒溫下培養(yǎng)24小時(shí),模擬沒有熱應(yīng)激的奶牛的體溫���。(b)來自早期竇卵泡(直徑0.5-1毫米)的OCGC在IVG培養(yǎng)中培養(yǎng)0���、4���、8或12天。在IVG培養(yǎng)的第0天評(píng)估卵母細(xì)胞直徑��。在IVG培養(yǎng)期間(第4���、8和12天)每4天評(píng)估OCGC的形態(tài)(OCGC的活力和顆粒細(xì)胞層中的腔形成)��。經(jīng)過12天的IVG���,一些存活的OCGC進(jìn)行了IVM����。在IVM之后,評(píng)估了一些卵母細(xì)胞的直徑和核狀態(tài)�����。IVM后的一些卵母細(xì)胞進(jìn)行IVF和IVC以評(píng)估發(fā)育能力�����。在IVM(生長和核狀態(tài)評(píng)估)或IVC(發(fā)育能力評(píng)估)之前培養(yǎng)的OCGC來自不同的培養(yǎng)階段��。在IVG培養(yǎng)期間,每4天評(píng)估IVG培養(yǎng)基中E2和P4的濃度��,及卵母細(xì)胞中的ROS和GSH水平(第4����、8和12天)。在第0天和IVG培養(yǎng)期間每4天(第4��、8和12天)評(píng)估TZP的數(shù)量���。
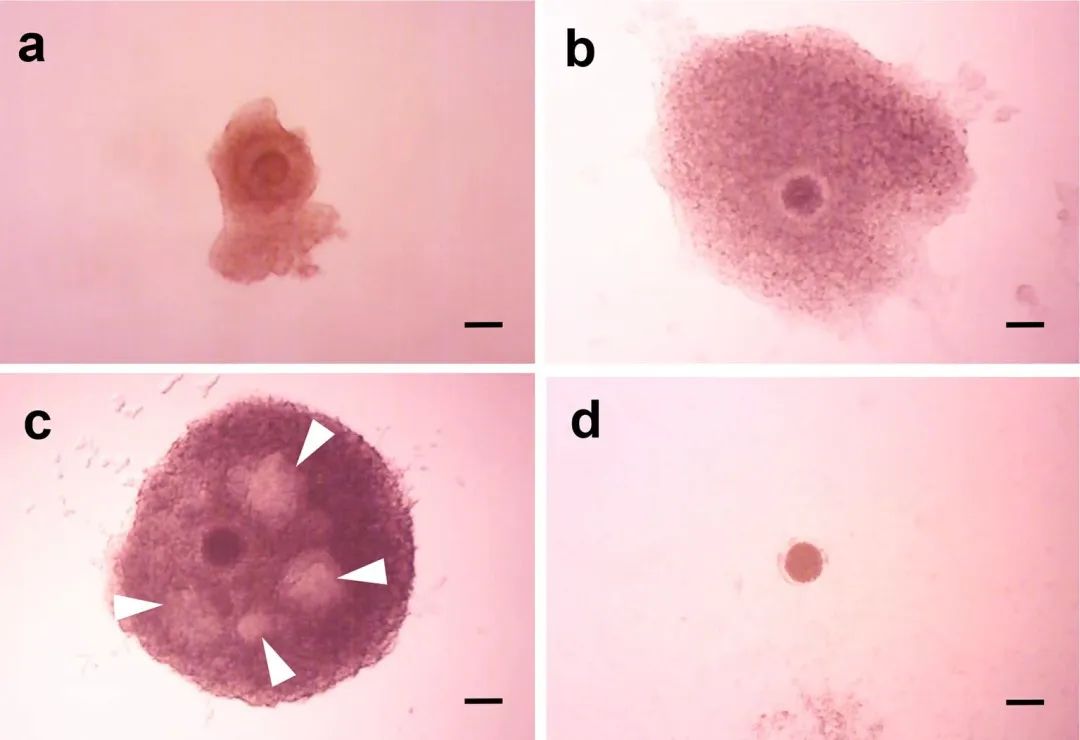
圖2.IVG培養(yǎng)12天前后OCGC的形態(tài)�。(a)IVG培養(yǎng)前分離的OCGC���。(b) IVG培養(yǎng)12天后�,在顆粒細(xì)胞層中沒有腔形成的OCGC存活��。(c)在顆粒細(xì)胞層中存活的OCGC形成腔(白色箭頭)�����。(d) IVG培養(yǎng)12天后退化的OCGC��。比例尺=100 μm。
圖3.IVG過程中熱休克對(duì)OCGC的活力(a)�����、腔形成的影響(b)�����,以及IVM后卵母細(xì)胞的生長和核狀態(tài)(c)��。括號(hào)中的數(shù)字表示OCGC的數(shù)量���。(a)OCGCs的存活率是根據(jù)689個(gè)OCGCs計(jì)算得出的���,這些OCGCs被培養(yǎng)到IVG培養(yǎng)結(jié)束(12天)(對(duì)照組345個(gè),熱休克組344個(gè))��。(b)根據(jù)第12天存活的406個(gè)OCGC(對(duì)照組208個(gè)�,熱休克組198個(gè))計(jì)算顆粒細(xì)胞層中腔形成的百分比�。abc不同字母表示同組培養(yǎng)時(shí)間差異顯著(P < 0.05)。(c)圖中方框上的線描繪了第25���、50和75個(gè)百分位數(shù)����,而虛線描繪了第10個(gè)和第90個(gè)百分位數(shù)。圖中方框下方的值表示卵母細(xì)胞的平均直徑(μm)�。括號(hào)中的數(shù)字表示為IVM的卵母細(xì)胞數(shù)量。a-c不同字母表示差異顯著(P < 0.01)���。
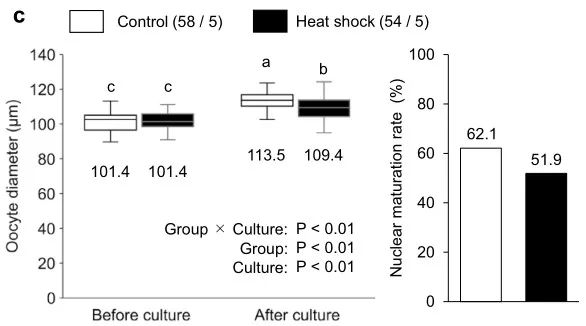
表1. OCGC體外生長培養(yǎng)過程中熱暴露對(duì)其卵母細(xì)胞發(fā)育能力的影響�。囊胚中的細(xì)胞數(shù)表示為平均值±SEM���。*囊胚率是根據(jù)受精卵母細(xì)胞的數(shù)量計(jì)算的�。不同上標(biāo)表示組間差異顯著(P < 0.05)��。
實(shí)驗(yàn)2:
IVG培養(yǎng)過程中熱暴露對(duì)顆粒細(xì)胞類固醇生成����、卵母細(xì)胞中ROS和GSH水平以及卵母細(xì)胞中TZP數(shù)量的影響
為了闡明熱休克組發(fā)育能力受損的機(jī)制,我們?cè)u(píng)估了顆粒細(xì)胞產(chǎn)生的E2和P4���,卵母細(xì)胞中的細(xì)胞內(nèi)ROS和GSH水平���,及卵母細(xì)胞和卵丘細(xì)胞間的TZP數(shù)量,這與卵母細(xì)胞的發(fā)育能力有關(guān)(圖1b)���。對(duì)照組和熱休克組的E2和P4生產(chǎn)水平相似(圖4)����。在兩組中,從第0-4天到第4-8天的E2量增加一直保持到IVG培養(yǎng)結(jié)束(第8-12天)��。另一方面���,P4含量在培養(yǎng)期間不斷增加���。兩組之間的E2/P4比率沒有差異。兩組卵母細(xì)胞中的細(xì)胞內(nèi)ROS水平?jīng)]有差異(圖5)�����;然而���,在第8天和第12天�,熱休克組卵母細(xì)胞中的GSH水平低于對(duì)照組(P < 0.05)(圖6)��。卵母細(xì)胞和周圍卵丘細(xì)胞之間的TZP數(shù)量在兩組之間沒有差異(圖 7)���。在兩組中,TZP的數(shù)量在IVG培養(yǎng)的第8天都減少了。
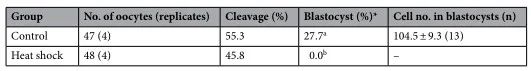
圖4. IVG培養(yǎng)過程中熱休克對(duì)OCGC產(chǎn)生E2和P4以及培養(yǎng)基中E2/P4比率的影響�����。不同字母表示同一組內(nèi)培養(yǎng)期之間存在顯著差異(P < 0.05)����。誤差線表示 SEM。
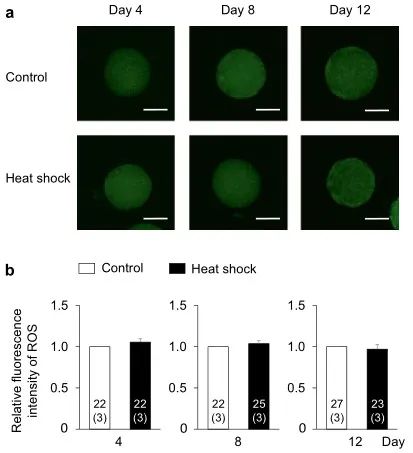
圖5. IVG培養(yǎng)過程中熱休克對(duì)卵母細(xì)胞細(xì)胞內(nèi)ROS水平的影響����。(a)用2',7'-二氯二氫熒光素二乙酸酯(DCHFDA)檢測的IVG卵母細(xì)胞的代表性熒光顯微照片�����。在對(duì)照組(上圖)和熱休克組(下圖)的IVG培養(yǎng)期間(第4�����、8和12 天)�����,每4天評(píng)估一次卵母細(xì)胞中的細(xì)胞內(nèi)ROS水平����。比例尺=50 μm��。(b)在IVG培養(yǎng)期間 (第4�、8和12天) 每4天對(duì)照和熱休克組的ROS水平的相對(duì)熒光強(qiáng)度��。使用總共141個(gè)卵母細(xì)胞(每個(gè)三個(gè)重復(fù))測量ROS的熒光強(qiáng)度����。條形圖中的數(shù)字表示卵母細(xì)胞的數(shù)量,而重復(fù)的數(shù)量顯示在括號(hào)中���。在同一培養(yǎng)日將熱休克組的熒光強(qiáng)度標(biāo)準(zhǔn)化為對(duì)照組的熒光強(qiáng)度����。

圖6. IVG培養(yǎng)過程中熱休克對(duì)卵母細(xì)胞內(nèi)GSH水平的影響����。(a)用CellTracker Blue檢測到的IVG卵母細(xì)胞的代表性熒光顯微照片。在對(duì)照組(上圖)和熱休克組(下圖)的IVG培養(yǎng)期間(第4��、8和12天)���,每4天評(píng)估一次卵母細(xì)胞中的細(xì)胞內(nèi)GSH水平����。比例尺=50 μm���。(b)在IVG培養(yǎng)期間(第4����、8和12天)��,對(duì)照組和熱休克組每4天的GSH水平的相對(duì)熒光強(qiáng)度����。使用總共177個(gè)卵母細(xì)胞(每個(gè)3-4個(gè)重復(fù))測量GSH的熒光強(qiáng)度。條形圖中的數(shù)字表示卵母細(xì)胞的數(shù)量�,而重復(fù)的數(shù)量顯示在括號(hào)中。在同一培養(yǎng)日將熱休克組的熒光強(qiáng)度標(biāo)準(zhǔn)化為對(duì)照組的熒光強(qiáng)度�����。*星號(hào)表示同一天對(duì)照組和熱休克組之間存在顯著差異(P < 0.05)�����。
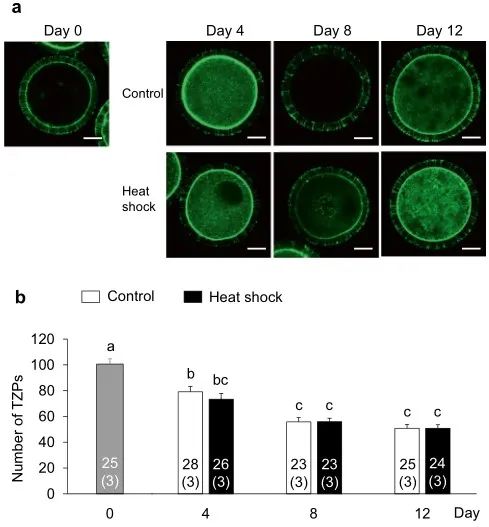
圖7. IVG培養(yǎng)過程中熱休克對(duì)卵母細(xì)胞和卵丘細(xì)胞之間的TZP數(shù)量的影響�����。(a)用熒光素異硫氰酸酯標(biāo)記的鬼筆環(huán)肽檢測卵母細(xì)胞和周圍卵丘細(xì)胞之間的TZP熒光染色。在對(duì)照組(上圖)和熱休克組(下圖)的第0天(左上圖)和IVG培養(yǎng)期間(第4��、8和12天)每4天評(píng)估TZP的數(shù)量����。比例尺=25μm。(b)在第0天����,及在IVG培養(yǎng)期間每4天(第4、8和12天)來自對(duì)照組和熱休克組的IVG卵母細(xì)胞中TZP的數(shù)量���。使用總共174個(gè)卵母細(xì)胞(每個(gè)三個(gè)重復(fù))評(píng)估TZP的數(shù)量��。條形圖中的數(shù)字表示卵母細(xì)胞的數(shù)量���,而重復(fù)的數(shù)量顯示在括號(hào)中。不同字母表示差異顯著(P < 0.05)��。
實(shí)驗(yàn)3:
補(bǔ)充半胱氨酸對(duì)IVG培養(yǎng)過程中高溫暴露的卵母細(xì)胞發(fā)育能力和GSH水平的影響
為了闡明促進(jìn)OCGC中的GSH合成是否能恢復(fù)IVF期間暴露于高溫的卵母細(xì)胞的發(fā)育能力�,我們研究了在培養(yǎng)基中補(bǔ)充半胱氨酸對(duì)IVF期間暴露于高溫的卵母細(xì)胞發(fā)育能力的影響。在實(shí)驗(yàn)中,我們檢查了未處理的半胱氨酸組(補(bǔ)充有0.0 mM半胱氨酸的IVG培養(yǎng)基)和半胱氨酸處理組(補(bǔ)充有1.2 mM半胱氨酸的IVG培養(yǎng)基)��。在之前的一項(xiàng)研究中���,向培養(yǎng)基中添加1.2 mM半胱氨酸可提高GSH水平并提高IVM期間暴露于高溫的卵母細(xì)胞的發(fā)育能力,因此本研究采用該半胱氨酸濃度���。半胱氨酸處理組(67.4%)和半胱氨酸未處理組(57.1%)之間的卵裂率相似(表2)����。然而����,半胱氨酸處理組的囊胚率(27.9%)顯著高于半胱氨酸未處理組(6.1%)(P < 0.05)。此外�����,半胱氨酸處理組的囊胚細(xì)胞數(shù)(122.1±10.8)略高于半胱氨酸未處理組(74.3±6.8)(P=0.06)���。IVG培養(yǎng)12天后卵母細(xì)胞中的細(xì)胞內(nèi)GSH水平在半胱氨酸處理組中高于半胱氨酸未處理組(P<0.05)(圖8)���。
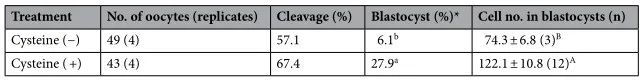
表2.半胱氨酸補(bǔ)充劑對(duì)IVG培養(yǎng)過程中暴露于高溫的卵母細(xì)胞發(fā)育能力的影響。囊胚中的細(xì)胞數(shù)表示為平均值±SEM�����。*囊胚率是根據(jù)受精卵母細(xì)胞的數(shù)量計(jì)算的。不同上標(biāo)表示組間差異顯著(P < 0.05)�����。A ,B不同的上標(biāo)表示組間的細(xì)微差異(P = 0.06)����。
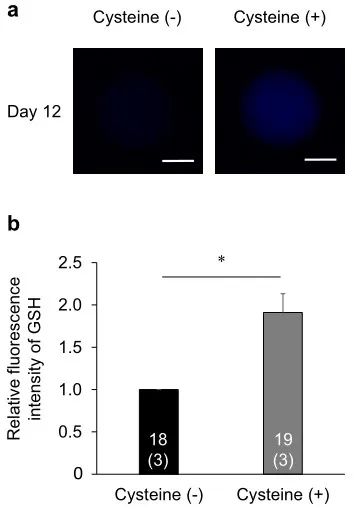
圖8.半胱氨酸補(bǔ)充劑對(duì)IVG培養(yǎng)過程中暴露于高溫的卵母細(xì)胞GSH水平的影響。(a)用CellTracker Blue檢測到的IVG卵母細(xì)胞的代表性熒光顯微照片����。在半胱氨酸未處理組(左圖)和半胱氨酸處理組(右圖)的IVG培養(yǎng)第12天評(píng)估細(xì)胞內(nèi)GSH水平。比例尺=50μm�����。(b) IVG培養(yǎng)第12天后半胱氨酸未處理組和處理組GSH水平的相對(duì)熒光強(qiáng)度�����。使用總共37個(gè)卵母細(xì)胞(每個(gè)重復(fù)三個(gè))測量GSH的熒光強(qiáng)度�����。條形圖中的數(shù)字表示卵母細(xì)胞的數(shù)量,而重復(fù)的數(shù)量顯示在括號(hào)中�����。半胱氨酸處理組中卵母細(xì)胞的熒光強(qiáng)度標(biāo)準(zhǔn)化為半胱氨酸未處理組中的那些�����。*星號(hào)表示半胱氨酸未處理組和處理組之間存在顯著差異(P < 0.05)����。
討論
據(jù)我們所知�����,我們首次證明�,IVG培養(yǎng)過程中的熱暴露會(huì)損害源自早期竇卵泡(0.5-1 mm)的卵母細(xì)胞的生長和發(fā)育能力。卵母細(xì)胞中的細(xì)胞內(nèi)GSH耗竭可能是早期竇狀卵泡的卵母細(xì)胞生長過程中熱暴露導(dǎo)致卵母細(xì)胞發(fā)育能力受損的原因之一��。
盡管對(duì)照組和熱休克組的卵裂率相似��,但熱休克組的囊胚率明顯低于對(duì)照組���。結(jié)果與之前的研究相似�,研究比較了夏季和冬季來自3至8毫米卵泡的卵母細(xì)胞的發(fā)育能力。研究還表明���,各組之間的卵裂率相似或夏季略低于冬季��,而夏季囊胚率明顯低于冬季���。然而,他們比較了兩個(gè)不同實(shí)驗(yàn)術(shù)語和使用不同卵母細(xì)胞來源的結(jié)果����。我們的實(shí)驗(yàn)?zāi)P涂捎糜谘芯肯募緹釕?yīng)激損害在對(duì)照組和熱休克組之間同時(shí)使用相同卵母細(xì)胞來源的卵母細(xì)胞發(fā)育能力的機(jī)制。
熱休克組IVG和IVM后卵母細(xì)胞的平均直徑明顯小于對(duì)照組�。卵母細(xì)胞生長受損可能是與熱暴露引起的卵母細(xì)胞發(fā)育能力降低相關(guān)的特征之一。眾所周知��,減數(shù)分裂成熟和隨后的胚胎發(fā)育的全部能力是在卵母細(xì)胞直徑約為110μm時(shí)獲得的����。因此,卵母細(xì)胞直徑≥110μm可能是確定卵母細(xì)胞是否會(huì)獲得后續(xù)發(fā)育能力的重要指標(biāo)�����。在本研究中,對(duì)照組(75.9%)的卵母細(xì)胞直徑≥110μm的百分比高于熱休克組(46.3%)(P<0.01)����,與之前的研究一致。
對(duì)照組和熱休克組之間的E2和P4生產(chǎn)水平?jīng)]有差異�。腔形成率(可以作為OCGC中健康類固醇生成的指標(biāo))在兩組之間沒有差異。先前的研究表明����,在牛IVG中,形成腔的OCGC比沒有腔的OCGC產(chǎn)生更多的E2和更少的P4�。另一方面,夏季熱應(yīng)激降低了泌乳后期奶牛外周E2濃度和優(yōu)勢(shì)卵泡液中E2濃度�。泌乳奶牛優(yōu)勢(shì)卵泡的卵泡液中P4濃度不受季節(jié)性熱應(yīng)激的影響�。與體內(nèi)情況相比,我們無法發(fā)現(xiàn)E2產(chǎn)生有任何差異的一個(gè)可能原因可能是本研究中IVG培養(yǎng)系統(tǒng)中沒有膜細(xì)胞��。熱應(yīng)激可能通過抑制全身內(nèi)分泌系統(tǒng)或體內(nèi)膜細(xì)胞的功能來抑制E2的產(chǎn)生���。先前顯示熱應(yīng)激可減少泌乳奶牛的促黃體激素(LH)數(shù)量���。此外,膜細(xì)胞對(duì)熱應(yīng)激高度敏感�����;季節(jié)性熱應(yīng)激大大降低了膜細(xì)胞A4的產(chǎn)生,并且這種減少似乎在夏季熱應(yīng)激結(jié)束后會(huì)持續(xù)很長時(shí)間��。這些變化可能導(dǎo)致體內(nèi)顆粒細(xì)胞分泌E2的下降���。研究結(jié)果清楚地表明��,來自早期竇卵泡的顆粒細(xì)胞對(duì)高溫的抵抗力相對(duì)較高����,以維持類固醇生成�����,這是卵泡生長過程中顆粒細(xì)胞的主要生理功能�����。
盡管對(duì)照組和熱休克組卵母細(xì)胞中的細(xì)胞內(nèi)ROS水平?jīng)]有差異�����,但熱休克組卵母細(xì)胞中的GSH水平低于對(duì)照組��。GSH是最重要的ROS清除劑之一。熱休克組的GSH消耗可能高于對(duì)照組�����,以減輕卵母細(xì)胞細(xì)胞質(zhì)中增加的氧化應(yīng)激��。在本研究中�,在第8天和第12天觀察到熱休克組的GSH水平下降。即使在來自早期竇卵泡的OCGC的牛IVG中����,卵母細(xì)胞的直徑也會(huì)顯著變大。此外����,在OCGC的IVG培養(yǎng)過程中,顆粒細(xì)胞的數(shù)量在第4天和第12天之間顯著增加���。因此,隨著培養(yǎng)時(shí)間的延長�����,對(duì)GSH合成的氨基酸(半胱氨酸��、谷氨酸、甘氨酸)的需求可能會(huì)增加����,以支持卵母細(xì)胞的生長、顆粒細(xì)胞的增殖�,并維持OCGCs的氧化應(yīng)激。因此�,在此期間從卵丘顆粒細(xì)胞到卵母細(xì)胞的GSH供應(yīng)可能在熱休克組中有所減少。雖然我們假設(shè)熱休克組中的GSH消耗也可能是由于生長中的卵母細(xì)胞與周圍卵丘細(xì)胞之間的細(xì)胞內(nèi)通訊受損引起的�,但對(duì)照組和熱休克組之間的TZP數(shù)量沒有差異。結(jié)果表明���,熱休克組中卵母細(xì)胞與周圍細(xì)胞之間的通訊可以維持在與對(duì)照組相同的水平��,而熱休克組中顆粒細(xì)胞或卵母細(xì)胞本身的谷胱甘肽產(chǎn)生減少�����。在未來的研究中���,我們應(yīng)該檢查OCGCs中與GSH合成和消耗相關(guān)的酶及其氨基酸代謝,這是IVG培養(yǎng)基中GSH合成所必需的��。
與對(duì)照組相比���,熱休克組的OCGC表現(xiàn)出較低的發(fā)育能力����,同時(shí)卵母細(xì)胞中的細(xì)胞內(nèi)GSH水平降低。此外�����,刺激GSH合成的半胱氨酸的補(bǔ)充增加了體外受精期間暴露于高溫的卵母細(xì)胞的細(xì)胞內(nèi)GSH水平和發(fā)育能力�����。表明IVM之前或之后卵母細(xì)胞中的低GSH水平與其發(fā)育能力受損之間存在關(guān)系���。IVM期間的熱暴露或在IVM前培養(yǎng)基中添加GSH合成抑制劑會(huì)降低卵母細(xì)胞中的細(xì)胞內(nèi)GSH水平及其發(fā)育能力�。這種受到細(xì)胞內(nèi)GSH耗竭導(dǎo)致的卵母細(xì)胞發(fā)育能力降低可能是由于隨后胚胎發(fā)育過程中抗氧化能力的降低��。目前的結(jié)果表明�����,在生長期的OCGC熱暴露會(huì)通過耗盡卵母細(xì)胞細(xì)胞質(zhì)中的GSH來損害發(fā)育能力�。
本研究表明����,來源于早期竇卵泡的OCGCs在生理范圍內(nèi)對(duì)高溫敏感��。早期竇卵泡(直徑0.5-1mm)發(fā)育成大的優(yōu)勢(shì)卵泡大約需要一個(gè)月的時(shí)間����;因此�����,夏季早期竇卵泡受熱導(dǎo)致的卵母細(xì)胞發(fā)育能力受損可能與隨后較冷的秋季生育能力低下有關(guān)�����。一些提高夏季抗氧化能力的治療或喂養(yǎng)管理可以改善熱應(yīng)激對(duì)早期竇卵泡的負(fù)面影響����,從而提高秋季的卵母細(xì)胞質(zhì)量和生育能力。本研究中開發(fā)的培養(yǎng)系統(tǒng)可以替代體內(nèi)試驗(yàn)�,以尋找可能的抗氧化劑來改善夏季熱應(yīng)激持續(xù)影響導(dǎo)致的低卵母細(xì)胞質(zhì)量和生育能力。
總之��,早期竇卵泡OCGC在IVG培養(yǎng)過程中的熱暴露會(huì)損害卵母細(xì)胞的生長和發(fā)育至囊胚期的卵母細(xì)胞百分比���。本研究表明��,卵母細(xì)胞內(nèi)GSH耗竭(抗氧化能力下降)是夏季熱應(yīng)激導(dǎo)致卵母細(xì)胞發(fā)育能力低下的原因之一����,這可能導(dǎo)致隨后秋季的生育能力受損。然而�����,我們?cè)诒狙芯恐嘘P(guān)注生理范圍內(nèi)的高溫對(duì)生長期培養(yǎng)的卵母細(xì)胞的影響����。體內(nèi)季節(jié)性熱應(yīng)激引起的GV卵母細(xì)胞發(fā)育能力低下不僅涉及體溫高,而且促性腺激素分泌和代謝系統(tǒng)的改變也將參與其中�。需要進(jìn)一步的研究來闡明夏季熱應(yīng)激在卵母細(xì)胞生長過程中降低卵母細(xì)胞能力的更詳細(xì)機(jī)制。
來源:Kohei Kawano, Kenichiro Sakaguchi�,et al. Effect of heat exposure on the growth and developmental competence of bovine oocytes derived from early antral follicles[J]. Scientific Reports ,2022, 12:8857
編譯:佳斌

